Вредные бактерии названия. Полезные и вредные бактерии. Какие бактерии самые опасные для человека
Размножение бактерий путем деления — самый распространенный метод увеличения численности микробной популяции. После деления происходит рост бактерий до исходного размера, для чего необходимы определенные вещества (факторы роста).
Способы размножения бактерий различны, но для большинства их видов присуща форма бесполового размножения способом деления. Способом почкования бактерии размножаются исключительно редко. Половое размножение бактерий присутствует в примитивной форме.
Рис. 1. На фото бактериальная клетка в стадии деления.
Генетический аппарат бактерий
Генетический аппарат бактерий представлен единственной ДНК — хромосомой. ДНК замкнута в кольцо. Хромосома локализована в нуклеотиде, не имеющем мембраны. В бактериальной клетке имеются плазмиды.
Нуклеоид
Нуклеоид является аналогом ядра. Он расположен в центре клетки. В нем локализована ДНК — носитель наследственной информации в свернутом виде. Раскрученная ДНК достигает в длину 1 мм. Ядерное вещество бактериальной клетки не имеет мембраны, ядрышка и набора хромосом, не делится митозом. Перед делением нуклеотид удваивается. Во время деления число нуклеотидов увеличивается до 4-х.

Рис. 2. На фото бактериальная клетка на срезе. В центральной части виден нуклеотид.
Плазмиды
Плазмиды представляют собой автономные молекулы свернутые в кольцо двунитевой ДНК. Их масса значительно меньше массы нуклеотида. Несмотря на то, что в ДНК плазмид закодирована наследственная информация, они не являются жизненно важными и необходимыми для бактериальной клетки.

Рис. 3. На фото бактериальная плазмида.
Этапы деления
После достижения определенных размеров, присущих взрослой клетке, запускаются механизмы деления.
Репликация ДНК
Репликация ДНК предшествует клеточному делению. Мезосомы (складки цитоплазматической мембраны) удерживают ДНК до тех пор, пока процесс деления (репликации) не завершится.
Репликация ДНК осуществляется с помощью ферментов ДНК-полимеразами. При репликации водородные связи в 2-х спиральной ДНК разрываются, в результате чего из одной ДНК образуются две дочерние односпиральные. В последующем, когда дочерние ДНК заняли свое место в разделенных дочерних клетках, происходит их восстановление.
Как только репликация ДНК завершилась, в результате синтеза появляется перетяжка, разделяющая клетку пополам. Вначале делению подвергается нуклеотид, затем цитоплазма. Синтез клеточной стенки завершает деление.
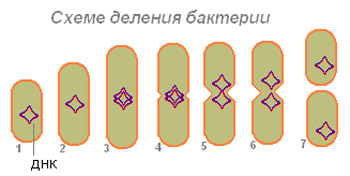
Рис. 4. Схема деления бактериальной клетки.
Обмен участками ДНК
У сенной палочки процесс репликации ДНК завершается обменом участками 2-х ДНК.
После деления клетки образуется перемычка, по которой ДНК одной клетки переходит в другую. Далее обе ДНК сплетаются. Некоторые отрезки обоих ДНК слипаются. В местах слипания происходит обмен отрезками ДНК. Одна из ДНК по перемычке уходит обратно в первую клетку.

Рис. 5. Вариант обмена ДНК у сенной палочки.
Типы делений бактериальных клеток
Если клеточное деление опережает процесс разделения, то образуются многоклеточные палочки и кокки.
При синхронном клеточном делении образуются две полноценные дочерние клетки.
Если нуклеотид делится быстрее самой клетки, то образуются многонуклеотидные бактерии.
Способы разделения бактерий
Деление с помощью разламывания
Деление с помощью разламывания характерно для сибиреязвенных бацилл. В результате такого деления клетки переламываются в местах сочленения, разрывая цитоплазматические мостики. Далее отталкиваются друг от друга, образуя цепочки.
Скользящее разделение
При скользящем разделении после деления клетка обосабливается и как бы скользит по поверхности другой клетки. Данный способ разделения характерен для некоторых форм эшерихий.
Секущееся разделение
При секущемся разделении одна из разделившихся клеток свободным концом описывает дугу круга, центром которого является точка ее контакта с другой клеткой, образуя римскую пятерку или клинопись (коринебактерии дифтерии, листерии).

Рис. 6. На фото бактерии палочковидной формы, образующие цепочки (сибиреязвенные палочки).

Рис. 7. На фото скользящий способ разделения кишечных палочек.

Рис. 8. Секущийся способ разделения коринебактерий.
Вид скоплений бактерий после деления
Скопления делящихся клеток имеют разнообразную форму, которая зависит от направления плоскости деления.
Шаровидные бактерии располагаются по одному, по двое (диплококки), пакетами, цепочками или как гроздья винограда. Палочковидные бактерии — цепочками.
Спиралевидные бактерии — хаотично.

Рис. 9. На фото микрококки. Они круглые, гладкие, имеют белую, желтую и красную окраску. В природе микрококки распространены повсеместно. Живут в разных полостях человеческого организма.

Рис. 10. На фото бактерии диплококки — Streptococcus pneumoniae.

Рис. 11. На фото бактерии сарцины. Кокковидные бактерии соединяются в пакеты.

Рис. 12. На фото бактерии стрептококки (от греческого «стрептос» — цепочка). Располагаются цепочками. Являются возбудителями целого ряда заболеваний.

Рис. 13. На фото бактерии «золотистые» стафилококки. Располагаются, как «гроздья винограда». Скопления имеют золотистую окраску. Являются возбудителями целого ряда заболеваний.

Рис. 14. На фото извитые бактерии лептоспиры — возбудители многих заболеваний.

Рис. 15. На фото палочковидные бактерии рода Vibrio.
Скорость деления бактерий
Скорость деления бактерий крайне высока. В среднем одна бактериальная клетка делится каждые 20 минут. В течение только одних суток одна клетка образует 72 поколения потомства. Микобактерии туберкулеза делятся медленно. Весь процесс деления занимает у них около 14 часов.

Рис. 16. На фото отображен процесс деления клетки стрептококка.
Половое размножение бактерий
В 1946 году учеными было обнаружено половое размножение в примитивной форме. При этом гаметы (мужские и женские половые клетки) не образуются, однако некоторые клетки обмениваются генетическим материалом (генетическая рекомбинация ).
Передача генов осуществляется в результате конъюгации — однонаправленного переноса части генетической информации в виде плазмид при контакте бактериальных клеток.
Плазмиды представляют собой молекулы ДНК небольшого размера. Они не связаны с геномом хромосом и способны удваиваться автономно. В плазмидах содержаться гены, которые повышают устойчивость бактериальных клеток к неблагоприятным условиям внешней среды. Бактерии часто передают эти гены друг другу. Отмечается так же передача генной информации бактериям другого вида.
При отсутствии истинного полового процесса именно конъюгация играет огромную роль при обмене полезными признаками. Так передается способность бактерий проявлять лекарственную устойчивость. Для человечества особо опасным является передача устойчивости к антибиотикам между болезнетворными популяциями.

Рис. 17. На фото момент конъюгации двух кишечных палочек.
Фазы развития бактериальной популяции
При посевах на питательную среду развитие бактериальной популяции проходит несколько фаз.
Исходная фаза
Исходная фаза — это период от момента посева до их роста. В среднем исходная фаза длится 1 — 2 часа.
Фаза задержки размножения
Это фаза интенсивного роста бактерий. Ее длительность составляет около 2-х часов. Она зависит от возраста культуры, периода приспособления, качества питательной среды и др.
Логарифмическая фаза
В эту фазу отмечается пик скорости размножения и увеличения бактериальной популяции. Ее длительность составляет 5 — 6 часов.
Фаза отрицательного ускорения
В эту фазу отмечается спад скорости размножения, уменьшается количество делящихся и увеличивается число погибших бактерий. Причина отрицательного ускорения — истощение питательной среды. Ее длительность составляет около 2-х часов.
Стационарная фаза максимума
В стационарную фазу отмечается равное количество погибших и вновь образованных особей. Ее длительность составляет около 2-х часов.
Фаза ускорения гибели
В эту фазу прогрессивно нарастает количество погибших клеток. Ее длительность составляет около 3-х часов.
Фаза логарифмической гибели
В эту фазу клетки бактерий отмирают с постоянной скоростью. Ее длительность составляет около 5-и часов.
Фаза уменьшения скорости отмирания
В эту фазу оставшиеся живыми клетки бактерий переходят в состояние покоя.

Рис. 18. На рисунке отображена кривая роста бактериальной популяции.

Рис. 19. На фото колонии синегнойной палочки сине-зеленого цвета, колонии микрококков желтого цвета, колонии Bacterium prodigiosum кроваво-красного цвета и колонии Bacteroides niger черного цвета.

Рис. 20. На фото колонии бактерий. Каждая колония — потомство одной-единственной клетки. В колонии число клеток исчисляется миллионами. вырастает колония за 1 — 3 суток.
Деление магниточувствительных бактерий
В 1970-х годах были открыты бактерии, обитающие в морях, которые обладали чувством магнетизма. Магнетизм позволяет этим удивительным существам ориентироваться по линиям магнитного поля Земли и находить серу, кислород и другие, так необходимые ей вещества. Их «компас» представлен магнитосомами, которые состоят из магнита. При делении магниточувствительные бактерии делят свой компас. При этом перетяжки при делении становится явно недостаточно, поэтому бактериальная клетка сгибается и делает резкий перелом.

Рис. 21. На фото момент деления магниточувствительной бактерии.
Рост бактерий
Вначале деления бактериальной клетки две молекулы ДНК расходятся в разные концы клетки. Далее клетка делится на две равноценные части, которые отделяются друг от друга и увеличиваются до исходного размера. Скорость деления многих бактерий составляет в среднем 20 — 30 минут. В течение только одних суток одна клетка образует 72 поколения потомства.
Масса клеток в процессе роста и развития быстро поглощает питательные вещества из окружающей среды. Этому способствуют благоприятные факторы внешней среды — температурный режим, достаточное количество питательных веществ, необходимая pH среды. Для клеток аэробов необходим кислород. Для анаэробов он представляет опасность. Однако безграничное размножение бактерий в природе не происходит. Солнечный свет, сухой воздух, недостаток пищи, высокая температура окружающей среды и другие факторы губительно действуют на бактериальную клетку.

Рис. 22. На фото момент деления клетки.
Факторы роста
Для роста бактерий необходимы определенные вещества (факторы роста), часть из которых синтезируется самой клеткой, часть поступает из окружающей среды. Потребность в факторах роста у всех бактерий разная.
Потребность в факторах роста является постоянным признаком, что позволяет использовать его для идентификации бактерий, подготовке питательных сред и использовать в биотехнологии.
Факторы роста бактерий (бактериальные витамины) — химические элементы, большинством из которых являются водорастворимые витамины группы В. В эту группу входят так же гемин, холин, пуриновые и пиримидиновые основания и другие аминокислоты. При отсутствии факторов роста наступает бактериостаз.
Бактерии используют факторы роста в минимальных количествах и в неизменном виде. Ряд химических веществ этой группы входят в состав клеточных ферментов.

Рис. 23. На фото момент деления палочковидной бактерии.
Важнейшие бактериальные факторы роста
- Витамин В1 (тиамин) . Принимает участие в углеводном обмене.
- Витамин В2» (рибофлавин) . Принимает участие в окислительно-восстановительных реакциях.
- Пантотеновая кислота является составной частью кофермента А.
- Витамин В6 (пиридоксин) . Принимает участие в обмене аминокислот.
- Витамины В12 (кобаламины — вещества, содержащие кобальт). Принимают активное участие в синтезе нуклеотидов.
- Фолиевая кислота . Некоторые ее производные входят в состав ферментов, катализирующих процессы синтеза пуриновых и пиримидиновых оснований, а также некоторых аминокислот.
- Биотин . Участвует в азотистом обмене, а также катализирует синтез ненасыщенных жирных кислот.
- Витамин РР (никотиновая кислота). Участвует в окислительно-восстановительных реакциях, образовании ферментов и обмене липидов и углеводов.
- Витамин Н (парааминобензойная кислота). Является фактором роста многих бактерий, в том числе населяющих кишечник человека. Из парааминобензойной кислоты синтезируется фолиевая кислота.
- Гемин . Является составной частью некоторых ферментов, которые принимают участие в реакциях окислениях.
- Холин . Принимает участие в реакциях синтеза липидов клеточной стенки. Является поставщиком метильной группы при синтезе аминокислот.
- Пуриновые и пиримидиновые основания (аденин, гуанин, ксантин, гипоксантин, цитозин, тимин и урацил). Вещества необходимы главным образом в качестве компонентов нуклеиновых кислот.
- Аминокислоты . Эти вещества являются составляющими белков клетки.
Потребность в факторах роста некоторых бактерий
Ауксотрофы для обеспечения жизнедеятельности нуждаются в поступлении химических веществ из вне. Например, клостридии не способны синтезировать лецитин и тирозин. Стафилококки нуждаются в поступлении лецитина и аргинина. Стрептококки нуждаются в поступлении жирных кислот — компонентов фосфолипидов. Коринебактерии и шигеллы нуждаются в поступлении никотиновой кислоты. Золотистые стафилококки, пневмококки и бруцеллы нуждаются в поступлении витамина В1. Стрептококки и бациллы столбняка — в пантотеновой кислоте.
Прототрофы самостоятельно синтезируют необходимые вещества.

Рис. 24. Разные условия окружающей среды по-разному влияют на рост колоний бактерий. Слева — стабильный рост в виде медленно расширяющегося круга. Справа — быстрый рост в виде «побегов».
Изучение потребности бактерий в факторах роста позволяет ученым получать большую микробную массу, так необходимую при изготовлении антимикробных препаратов, сывороток и вакцин.
Подробно о бактерияx читай в статьях:
Размножение бактерий является механизмом повышения числа микробной популяции. Деление бактерий — основной способ размножения. После деления бактерии должны достигнуть размеров взрослых особей. Рост бактерий происходит путем быстрого поглощения питательных веществ их окружающей среды. Для роста необходимы определенные вещества (факторы роста), часть из которых синтезирует сама бактериальная клетка, часть поступает из окружающей среды.
Бактерии это самый древний организм на земле, а также самый простой в своем строении. Он состоит всего из одной клетки, которую можно увидеть и изучить только под микроскопом. Характерным признаком бактерий является отсутствие ядра, вот почему бактерии относят к прокариотам.
Некоторые виды образовывают небольшие группы клеток, такие скопления могут быть окружены капсулой (чехлом). Размер, форма и цвет бактерии сильно зависит от окружающей среды.
По форме бактерии различаются на: палочковидные (бациллы), сферические (кокки) и извитые (спириллы). Встречаются и видоизмененные – кубические, С-образные, звездчатые. Их размеры колеблются от 1 до 10мкм. Отдельные виды бактерий могут активно передвигаться при помощи жгутиков. Последние иногда превышают размер самой бактерии в два раза.
Виды форм бактерий
Для движения бактерии используют жгутики, количество которых бывает различное – один, пара, пучок жгутиков. Расположение жгутиков также бывает разным – с одной стороны клетки, по бокам или равномерно распределены по всей плоскости. Также одним из способов передвижения считается скольжение благодаря слизи, которой покрыт прокариот. У большинства внутри цитоплазмы есть вакуоли. Регулировка ёмкости газа в вакуолях помогает им двигаться в жидкости вверх или вниз, а также перемещаться по воздушных каналах почвы.
Ученые открыли более 10 тысяч разновидностей бактерий, но по предположениям научных исследователей в мире существует их более миллиона видов. Общая характеристика бактерий дает возможность определиться с их ролью в биосфере, а также изучить строение, виды и классификацию царства бактерий.
Места обитания
Простота строения и быстрота адаптации к окружающим условиям помогла бактериям распространиться в широком диапазоне нашей планеты. Они существуют везде: вода, почва, воздух, живые организмы – всё это максимально приемлемое место обитания для прокариотов.
Бактерии находили как на южном полюсе, так и в гейзерах. Они есть на океанском дне, а также в верхних слоях воздушной оболочки Земли. Бактерии живут везде, но их количество зависит от благоприятных условий. К примеру, большая численность видов бактерий проживает в открытых водоемах, а также почве.
Особенности строения
Клетка бактерии отличается не только тем, что в ней нет ядра, но и отсутствием митохондрий и пластид. ДНК данного прокариота находится в специальной ядерной зоне и имеет вид замкнутого в кольцо нуклеоида. У бактерии строение клетки состоит из клеточной стенки, капсулы, капсулоподобной оболочки, жгутиков, пили и цитоплазматичной мембраны. Внутреннее строение оформляют цитоплазма, гранулы, мезосомы, рибосомы, плазмиды, включения и нуклеоид.

Клеточная стенка бактерии выполняет функцию обороны и опоры. Вещества могут свободно протекать сквозь неё, благодаря проницаемости. Данная оболочка имеет в своем составе пектин и гемицеллюлозу. Некоторые бактерии выделяют особую слизь, которая может помочь защититься от пересыхания. Слизь формирует капсулу – полисахарид по химическому составу. В такой форме бактерия способна переносить даже очень большие температуры. Также она выполняет и другие функции, к примеру слипание с любыми поверхностями.
На поверхности клетки бактерии находятся тонкие белковые ворсинки – пили. Их может быть большая численность. Пили помогают клетке передавать генетический материал, а также обеспечивают слипание с другими клетками.
Под плоскостью стенки находится трехслойная цитоплазматичная мембрана. Она гарантирует транспорт веществ, а также имеет немалую роль в образовании спор.
Цитоплазма бактерий на 75 процентов произведена из воды. Состав цитоплазмы:
- Рыбосомы;
- мезосомы;
- аминокислоты;
- ферменты;
- пигменты;
- сахар;
- гранулы и включения;
- нуклеоид.
Обмен веществ у прокариотов возможен, как с участием кислорода, так и без его него. Большая их часть питаются уже готовыми питательными веществами органического происхождения. Очень мало видов способны сами синтезировать органические вещества из неорганических. Это сине-зеленые бактерии и цианобактерии, которые отыграли немалую роль в формировании атмосферы и насыщении её кислородом.
Размножение
В условиях, благоприятных для размножения, оно осуществляется почкованием или вегетативно. Бесполое размножение происходит в такой последовательности:
- Клетка бактерии достигает максимального объема и содержит необходимый запас питательных веществ.
- Клетка удлиняется, посередине появляется перегородка.
- Внутри клетки происходит дележ нуклеотида.
- ДНК основная и отделенная расходятся.
- Клетка делится пополам.
- Остаточное формирование дочерних клеток.
При таком способе размножения нету обмена генетической информацией, поэтому все дочерние клетки будут точной копией материнской.
Процесс размножения бактерий в неблагоприятных условиях более интересен. О способности полового размножения бактерий ученые узнали сравнительно недавно – в 1946 году. У бактерий нет разделения на женские и половые клетки. Но ДНК у них встречается разнополое. Две такие клетки при приближении друг к другу образовывают канал для передачи ДНК, происходит обмен участками – рекомбинация. Процесс довольно длительный, результатом которого являются две совершенно новые особи.
Большинство бактерий очень сложно увидеть под микроскопом, так как они не имеют своей окраски. Немногие разновидности имеют пурпурный или зеленый окрас, благодаря содержанию в них бактериохлорофилла и бактериопурпурина. Хотя если рассматривать некоторые колонии бактерий, становится ясно, что они выделяют окрашиваемые вещества в среду обитания и приобретают яркую окраску. Для того, чтобы подробней изучать прокариотов, их окрашивают.

Классификация
Классификация бактерий может быть основана на таких показателях, как:
- Форма
- способ передвижения;
- способ получения энергии;
- продукты жизнедеятельности;
- степень опасности.

Бактерии симбионты живут в содружестве с иными организмами.
Бактерии сапрофиты проживают на уже отмерших организмах, продуктах и органических отходах. Они способствуют процессам гниения и брожения.
Гниение очищает природу от трупов и других отходов органического происхождения. Без процесса гниения не было бы круговорота веществ в природе. Так в чем же состоит роль бактерий в круговороте веществ?
Бактерии гниения - это помощник в процессе расщепления белковых соединений, а также жиров и других соединений, содержащих в себе азот. Проведя сложную химическую реакцию, они разрывают связи между молекулами органических организмов и захватывают молекулы белка, аминокислот. Расщепляясь, молекулы высвобождают аммиак, сероводород и другие вредные вещества. Они ядовиты и могут вызывать отравление у людей и животных.
Бактерии гниения быстро размножаются в благоприятных для них условиях. Так как это не только полезные бактерии, но и вредные, то чтобы не допустить преждевременного гниения у продуктов, люди научились их обрабатывать: сушить, мариновать, солить, коптить. Все эти способы обработки убивают бактерии и не дают им размножаться.
Бактерии брожения при помощи ферментов способны расщеплять углеводы. Эту способность люди заметили еще в древние времена и используют такие бактерии для изготовления молочнокислых продуктов, уксусов, а также других продуктов питания до сих пор.
Бактерии, трудясь в совокупности с другими организмами, делают очень важную химическую работу. Очень важно знать какие есть виды бактерий и какую пользу или вред приносят для природы.
Значение в природе и для человека
Выше уже отмечалось большое значение многих видов бактерий (при процессах гниения и различных типах брожения), т.е. выполнение санитарной роли на Земле.
Бактерии также играют огромную роль в круговороте углерода, кислорода, водорода, азота, фосфора, серы, кальция и других элементов. Многие виды бактерий способствуют активной фиксации атмосферного азота и переводят его в органическую форму, способствуя повышению плодородия почв. Особо важное значение имеют те бактерии, которые разлагают целлюлозу, являющиеся основным источником углерода для жизнедеятельности почвенных микроорганизмов.
Сульфатредуцирующие бактерии участвуют в образовании нефти и сероводорода в лечебных грязях, почвах и морях. Так, насыщенный сероводородом слой воды в Черном море является результатом жизнедеятельности сульфатредуцирующих бактерий. Деятельность этих бактерий в почвах приводит к образованию соды и содового засоления почвы. Сульфатредуцирующие бактерии переводят питательные вещества в почвах рисовых плантаций в такую форму, которая становится доступной для корней этой культуры. Эти бактерии могут вызывать коррозию металлических подземных и подводных сооружений.
Благодаря жизнедеятельности бактерий почва освобождается от многих продуктов и вредных организмов и насыщается ценными питательными веществами. Бактерицидные препараты успешно используются для борьбы с многими видами насекомых-вредителей (кукурузным мотыльком и др.).
Многие виды бактерий используются в различных отраслях промышленности для получения ацетона, этилового и бутилового спиртов, уксусной кислоты, ферментов, гормонов, витаминов, антибиотиков, белково-витаминных препаратов и т.д.
Без бактерий невозможны процессы при дублении кожи, сушке листьев табака, выработке шелка, каучука, обработке какао, кофе, мочении конопли, льна и других лубоволокнистых растений, квашении капусты, очистке сточных вод, выщелачивании металлов и т.д.
Морфология бактерий, строение прокариотической клетки.
В прокариотических клетках нет четкой границы между ядром и цитоплазмой, отсутствует ядерная мембрана. ДНК в этих клетках не образует структур, похожих на хромосомы эукариот. Поэтому у прокариот не происходят процессы митоза и мейоза. Большинство прокариот не образует внутриклеточных органелл, ограниченных мембранами. Кроме того, в прокариотических клетках нет митохондрий и хлоропластов.
Бактерии , как правило, являются одноклеточными организмами, клетка их имеет довольно простую форму, представляет собой шар или цилиндр, иногда изогнутый. Размножаются бактерии преимущественно делением на две равноценные клетки.
Бактерии шаровидной формы называются кокками и могут быть сферическими, эллипсоидальными, бобовидными и ланцетовидными.
По расположению клеток относительно друг друга после деления кокки подразделяют на несколько форм. Если после деления клетки расходятся и располагаются поодиночке, то такие формы называют монококками . Иногда кокки при делении образуют скоплений, напоминающие виноградную гроздь. Подобные формы относятся к стафилококкам . Кокки, остающиеся после деления в одной плоскости связанными парами, называются диплококками , а образующие различной длины цепочки - стрептококками . Сочетания из четырех кокков, появляющиеся после деления клетки в двух взаимно перпендикулярных плоскостях, представляют собой тетракокки . Некоторые кокки делятся в трех взаимно перпендикулярных плоскостях, что приводит к образованию своеобразных скоплений кубической формы, называемых сардинами.
Большинство бактерий имеют цилиндрическую , или палочковидную, форму. Палочковидные формы бактерий, образующие споры, именуют бациллами , а не образующие споры - бактериями .
Палочковидные бактерии различаются по форме, размеру в длину и в поперечнике, форме концов клетки, а также по взаимному расположению. Они могут иметь цилиндрическую форму с прямыми концами или овальную - с закругленными или заостренными концами. Бактерии бывают также слегка изогнутыми, встречаются нитевидные и ветвящиеся формы (например, микобактерии и актиномицеты).
В зависимости от взаимного расположения отдельных клеток после деления палочковидные бактерии делят на собственно палочки (одиночное расположение клеток), диплобактерии или диплобациллы (парное расположение клеток), стрептобактерии или стрептобациллы (образуют цепочки различной длины). Нередко встречаются извитые, или спиралевидные, бактерии. К этой группе относятся спириллы (от лат. spira - завиток), имеющие форму длинных изогнутых (от 4 до 6 витков) палочек, и вибрионы (лат. vibrio - изгибаюсь), представляющие собой лишь 1/4 часть витка спирали, похожие на запятую.
Известны нитевидные формы бактерий, обитающие в водоемах. Кроме перечисленных, встречаются многоклеточные бактерии, несущие на поверхности клетки протоплазм этические выросты - простеки, треугольные и звездообразные бактерии, а также имеющие форму замкнутого и незамкнутого кольца и червеобразные бактерии.
Клетки бактерий очень малы. Их измеряют в микрометрах, а детали тонкой структуры - в нанометрах. Кокки обычно имеют диаметр около 0,5-1,5 мкм. Ширина палочковидных (цилиндрических) форм бактерий в большинстве случаев колеблется от 0,5 до 1 мкм, а длина равняется нескольким микрометрам (2-10). Мелкие палочки имеют ширину 0,2-0,4 и длину 0,7-1,5 мкм. Среди бактерий могут встречаться и настоящие гиганты, длина которых достигает десятков и даже сотен микрометров. Формы и размеры бактерий значительно изменяются в зависимости от возраста культуры, состава среды и ее осмотических свойств, температуры и других факторов.
Из трех основных форм бактерий кокки наиболее стабильны по размерам, палочковидные бактерии более изменчивы, причем особенно значительно меняется длина клеток.
Бактериальная клетка, помещенная на поверхность твердой питательной среды, растет, делится, образуя колонию бактерий-потомков. Через несколько часов роста колония состоит уже из такого большого числа клеток, что ее можно видеть невооруженным глазом. Колонии могут иметь слизистую или пастообразную консистенцию, в некоторых случаях они бывают пигментированы. Иногда внешний вид колоний настолько характерен, что позволяет без особых трудностей провести идентификацию микроорганизмов.
Основы физиологии бактерий.
По химическому составу микроорганизмы мало отличаются от других живых клеток.
Вода составляет 75-85% , в ней растворены химические вещества.
Сухое вещество 15-25%, в состав входят органические и минеральные соединения
Питание бактерий. Поступление в бактериальную клетку питательных веществ осуществляется несколькими способами и зависит от концентрации веществ, величины молекул, рН среды, проницаемости мембран и др. По типу питания микроорганизмы делятся на:
автотрофы – синтезируют все углеродсодержащие вещества из СО2;
гетеротрофы – в качестве источника углерода используют органические вещества;
сапрофиты – питаются органическими веществами отмерших организмов;
Дыхание бактерий . Дыхание, или биологическое окисление основано на окислительно-восстановительных реакциях, идущих с образованием молекулы АТФ. По отношению к молекулярному кислороду бактерии можно разделить на три основные группы:
облигатные аэробы – могут расти только при наличии кислорода;
облигатные анаэробы – растут на среде без кислорода, который для них токсичен;
факультативные анаэробы – могут расти как при кислороде, так и без него.
Рост и размножение бактерий. Большинство прокариот размножаются бинарным делением пополам, реже почкованием и фрагментацией. Бактерии, как правило, характеризуются высокой скоростью размножения. Время деления клетки у различных бактерий колеблется довольно в широких пределах: от 20 минут у кишечной палочки до 14 часов у микобактерий туберкулеза. На плотных питательных средах бактерии образуют скопления клеток, называемые колониями.
Ферменты бактерий. Важную роль в обмене веществ микроорганизмов играют ферменты. Различают:
эндоферменты – локализуются в цитоплазме клеток;
экзоферменты – выделяются в окружающую среду.
Ферменты агрессии разрушают ткань и клетки, обусловливая широкое распространение микробов и их токсинов в инфицированной ткани. Биохимические свойства бактерий определяются составом ферментов:
сахаролитические –расщепление углеводов;
протеолитические – расщепление белков,
липолитические – расщепление жиров,
и являются важным диагностическим признаком при идентификации микроорганизмов.
Для многих патогенных микроорганизмов оптимальными являются температура 37°С и рН 7,2-7,4.
Вода. Значимость воды для бактерий. Вода составляет около 80% массы бактерий. Рост и развитие бактерий облигатно зависят от наличия воды, так как все химические реакции, протекающие в живых организмах, реализуются в водной среде. Для нормального роста и развития микроорганизмов необходимо присутствие воды в окружающей среде.
Для бактерий содержание воды в субстрате должно быть более 20%. Вода должна находиться в доступной форме: в жидкой фазе в интервале температур от 2 до 60 °С; этот интервал известен как биокинетическая зона. Хотя в химическом отношении вода весьма устойчива, продукты её ионизации - ионы Н+ и ОН" оказывают очень большое влияние на свойства практически всех компонентов клетки (белков, нуклеиновых кислит, липидов и т.д.). Так, каталитическая активность ферментов в значительной мере зависит от концентрации ионов Н+ и ОН".
Брожение как основной способ получения энергии у бактерий.
Брожение - это метаболический процесс, в результате которого образуется АТФ, а доноры и акцепторы электронов, это продукты образующиеся в ходе самого брожения.
Брожение – процесс ферментативного расщепления органических веществ, преимущественно углеводов, протекающий без использования кислорода. Служит источником энергии для жизнедеятельности организма и играет большую роль в круговороте веществ и в природе. Некоторые виды брожения, вызываемые микроорганизмами (спиртовое, молочнокислое, маслянокислое, уксуснокислое), используются в производстве этилового спирта, глицерина и других технических и пищевых продуктов.
Спиртовое брожение (осуществляется дрожжами и некоторыми видами бактерий), в ходе него пируват расщепляется на этанол и диоксид углерода. Из одной молекулы глюкозы в результате получается две молекулы спирта (этанола) и две молекулы углекислого газа. Этот вид брожения очень важен в производстве хлеба, пивоварении, виноделии и винокурении.
Молочнокислое брожение , в ходе которого пируват восстанавливается до молочной кислоты, осуществляют молочнокислые бактерии и другие организмы. При сбраживании молока молочнокислые бактерии преобразуют лактозу в молочную кислоту, превращая молоко в кисломолочные продукты (йогурт, простокваша и др.); молочная кислота придаёт этим продуктам кисловатый вкус.
Молочнокислое брожение происходит также в мышцах животных, когда потребность в энергии выше, чем обеспечиваемая дыханием, и кровь не успевает доставлять кислород.
Обжигающие ощущения в мышцах во время тяжелых физических упражнений соотносятся с получением молочной кислоты и сдвигом к анаэробному гликолизу, поскольку кислород преобразуется в диоксид углерода аэробным гликолизом быстрее, чем организм восполняет запас кислорода; а болезненность в мышцах после физических упражнений вызвана микротравмами мышечных волокон. Организм переходит к этому менее эффективному, но более скоростному методу производства АТФ в условиях недостатка кислорода. Затем печень избавляется от излишнего лактата, преобразуя его обратно в важное промежуточное звено гликолиза - пируват.
Уксуснокислое брожение осуществляют многие бактерии. Уксус (уксусная кислота) - прямой результат бактериальной ферментации. При мариновании продуктов уксусная кислота предохраняет пищу от болезнетворных и вызывающих гниение бактерий.
Маслянокислое брожение приводит к образованию масляной кислоты; его возбудителями являются некоторые анаэробные бактерии рода Клостридиум.
Размножение бактерий.
Некоторые бактерии не имеют полового процесса и размножаются лишь равновеликим бинарным поперечным делением или почкованием. Для одной группы одноклеточных цианобактерий описано множественное деление (ряд быстрых последовательных бинарных делений, приводящий к образованию от 4 до 1024 новых клеток). Для обеспечения необходимой для эволюции и приспособления к изменчивой окружающей среде пластичности генотипа у них существуют иные механизмы.
При делении большинство грамположительных бактерий и нитчатых цианобактерий синтезируют поперечную перегородку от периферии к центру при участии мезосом. Грамотрицательные бактерии делятся путём перетяжки: на месте деления обнаруживается постепенно увеличивающееся искривление ЦПМ и клеточной стенки внутрь. При почковании на одном из полюсов материнской клетки формируется и растёт почка, материнская клетка проявляет признаки старения и обычно не может дать более 4 дочерних. Почкование имеется у разных групп бактерий и, предположительно, возникало несколько раз в процессе эволюции.
У других бактерий кроме размножения наблюдается половой процесс, но в самой примитивной форме. Половой процесс бактерий отличается от полового процесса эукариот тем, что у бактерий не образуются гаметы и не происходит слияния клеток. Механизм рекомбинации у прокариот. Однако главнейшее событие полового процесса, а именно обмен генетическим материалом, происходит и в этом случае. Это называется генетической рекомбинацией. Часть ДНК (очень редко вся ДНК) клетки-донора переносится в клетку-реципиент, ДНК которой генетически отличается от ДНК донора. При этом перенесённая ДНК замещает часть ДНК реципиента. В процессе замещения ДНК участвуют ферменты, расщепляющие и вновь соединяющие цепи ДНК. При этом образуется ДНК, которая содержит гены обеих родительских клеток. Такую ДНК называют рекомбинантной. У потомства, или рекомбинантов, наблюдается заметное разнообразие признаков, вызванное смещением генов. Такое разнообразие признаков очень важно для эволюции и является главным преимуществом полового процесса.
Известны 3 способа получения рекомбинантов. Это - в порядке их открытия - трансформация, конъюгация и трансдукция.
Происхождение бактерий.
Бактерии наряду с археями были одними из первых живых организмов на Земле, появившись около 3,9-3,5 млрд лет назад. Эволюционные взаимоотношения между этими группами ещё до конца не изучены, есть как минимум три основные гипотезы: Н. Пэйс предполагает наличие у них общего предка протобактерии, Заварзин считает архей тупиковой ветвью эволюции эубактерий, освоившей экстремальные местообитания; наконец, по третьей гипотезе археи - первые живые организмы, от которых произошли бактерии.
Эукариоты возникли в результате симбиогенеза из бактериальных клеток намного позже: около 1,9-1,3 млрд лет назад. Для эволюции бактерий характерен ярко выраженный физиолого-биохимический уклон: при относительной бедности жизненных форм и примитивном строении, они освоили практически все известные сейчас биохимические процессы. Прокариотная биосфера имела уже все существующие сейчас пути трансформации вещества. Эукариоты, внедрившись в неё, изменили лишь количественные аспекты их функционирования, но не качественные, на многих этапах циклов элементов бактерии по-прежнему сохраняют монопольное положение.
Одними из древнейших бактерий являются цианобактерии. В породах, образованных 3,5 млрд лет назад, обнаружены продукты их жизнедеятельности - строматолиты, бесспорные свидетельства существования цианобактерий относятся ко времени 2,2-2,0 млрд лет назад. Благодаря им в атмосфере начал накапливаться кислород, который 2 млрд лет назад достиг концентраций, достаточных для начала аэробного дыхания. К этому времени относятся образования, свойственные облигатно аэробной Metallogenium.
Появление кислорода в атмосфере (кислородная катастрофа) нанесло серьёзный удар по анаэробным бактериям. Они либо вымирают, либо уходят в локально сохранившиеся бескислородные зоны. Общее видовое разнообразие бактерий в это время сокращается.
Предполагается, что из-за отсутствия полового процесса, эволюция бактерий идёт по совершенно иному механизму, нежели у эукариот. Постоянный горизонтальный перенос генов приводит к неоднозначностям в картине эволюционных связей, эволюция протекает крайне медленно (а, возможно, с появлением эукариот и вовсе прекратилась), зато в изменяющихся условиях происходит быстрое перераспределение генов между клетками при неизменном общем генетическом пуле.
Систематика бактерий.
Роль бактерий в природе и в жизни человека.
Бактерии играют важную роль на Земле. Они принимают самое активное участие в круговороте веществ в природе. Все органические соединения и значительная часть неорганических подвергаются с помощью бактерий существенным изменениям. Эта их роль в природе имеет глобальное значение. Появившись на Земле раньше всех организмов (более 3,5 млрд лет назад), они создали живую оболочку Земли и продолжают активно перерабатывать живое и мертвое органическое вещество, вовлекая продукты своего обмена в круговорот веществ. Круговорот веществ в природе является основой существования жизни на Земле.
Распад всех растительных и животных остатков и образование перегноя и гумуса тоже производится в основном бактериями. Бактерии – мощный биотический фактор в природе.
Огромное значение имеет почвообразовательная работа бактерий. Первая почва на нашей планете была создана бактериями. Однако и в наше время состояние и качество почвы зависят от функционирования почвенных бактерий. Особенно важны для плодородия почвы так называемые азотфиксирующие клубеньковые бактерии-симбионты бобовых растений. Они насыщают почву ценными азотными соединениями.
Бактерии очищают грязные сточные воды, расщепляя органические вещества и превращая их в безвредные неорганические. Это свойство бактерий широко используется в работе очистных сооружений.
Во многих случаях бактерии могут быть и вредны для человека. Так, сапротрофные бактерии портят пищевые продукты. Чтобы уберечь продукты от порчи, их подвергают специальной обработке (кипячение, стерилизация, замораживание, высушивание, химическая очистка и т. д.). Если этого не делать, могут произойти пищевые отравления.
Среди бактерий имеется много болезнетворных (патогенных) видов, вызывающих заболевания у людей, животных или растений. Тяжелое заболевание брюшной тиф вызывает бактерия сальмонелла, дизентерию – бактерия шигелла. Болезнетворные бактерии разносятся по воздуху с капельками слюны больного человека при чихании, кашле и даже при обычном разговоре (дифтерия, коклюш). Некоторые болезнетворные бактерии очень устойчивы к высыханию и долго сохраняются в пыли (туберкулезная палочка). В пыли и почве живут бактерии рода клостридиум – возбудители газовой гангрены и столбняка. Некоторые бактериальные заболевания передаются при физическом контакте с больным человеком (венерические болезни, проказа). Часто болезнетворные бактерии передаются человеку с помощью так называемых переносчиков. Например, мухи, ползая по нечистотам, собирают на своих лапках тысячи болезнетворных бактерий, а затем оставляют их на продуктах, потребляемых человеком.
История изучения
Основы общей микробиологии и изучения роли бактерий в природе заложили Бейеринк Мартинус Виллем и Виноградский Сергей Николаевич .
Изучение строения бактериальной клетки началось с изобретением электронного микроскопа в 1930-е годы . В 1937 году Э. Чаттон предложил делить все организмы по типу клеточного строения на прокариот и эукариот, и в 1961 году Стейниер и Ван Ниль окончательно оформили это разделение. Развитие молекулярной биологии привело к открытию в 1977 году К. Вёзе коренных различий и среди самих прокариот: между бактериями и археями.
Строение
Подавляющее большинство бактерий (за исключением актиномицетов и нитчатых цианобактерий) одноклеточны . По форме клеток они могут быть округлыми (кокки), палочковидными (бациллы , клостридии , псевдомонады), извитыми (вибрионы , спириллы , спирохеты), реже - звёздчатыми, тетраэдрическими , кубическими, C- или O-образными. Формой определяются такие способности бактерий, как прикрепление к поверхности, подвижность, поглощение питательных веществ. Отмечено, например, что олиготрофы , то есть бактерии, живущие при низком содержании питательных веществ в среде, стремятся увеличить отношение поверхности к объёму, например, с помощью образования выростов (т. н. простек).
Из обязательных клеточных структур выделяют три:
С внешней стороны от ЦПМ находятся несколько слоёв (клеточная стенка , капсула , слизистый чехол), называемых клеточной оболочкой , а также поверхностные структуры (жгутики , ворсинки). ЦПМ и цитоплазму объединяют вместе в понятие протопласт .
Строение протопласта
ЦПМ ограничивает содержимое клетки (цитоплазму) от внешней среды. Гомогенная фракция цитоплазмы, содержащая набор растворимых РНК , белков , продуктов и субстратов метаболических реакций, названа цитозолем . Другая часть цитоплазмы представлена различными структурными элементами.
Вся необходимая для жизнедеятельности бактерий генетическая информация содержится в одной ДНК (бактериальная хромосома), чаще всего имеющей форму ковалентно замкнутого кольца (линейные хромосомы обнаружены у Streptomyces и Borrelia ). Она в одной точке прикреплена к ЦПМ и помещается в структуре, обособленной, но не отделённой мембраной от цитоплазмы, и называемой нуклеоид . ДНК в развёрнутом состоянии имеет длину более 1 мм. Бактериальная хромосома представлена обычно в единственном экземпляре, то есть практически все прокариоты гаплоидны , хотя в определённых условиях одна клетка может содержать несколько копий своей хромосомы, а Burkholderia cepacia имеет три разных кольцевых хромосомы (длиной 3,6; 3,2 и 1,1 млн пар нуклеотидов). Рибосомы прокариот также отличны от таковых у эукариот и имеют константу седиментации 70 S (80 S у эукариот).
Помимо этих структур, в цитоплазме также могут находиться включения запасных веществ.
Клеточная оболочка и поверхностные структуры
У бактерий существует два основных типа строения клеточной стенки, свойственных грамположительным и грамотрицательным видам.
Клеточная стенка грамположительных бактерий представляет собой гомогенный слой толщиной 20-80 нм, построенный в основном из пептидогликана с меньшим количеством тейхоевых кислот и небольшим количеством полисахаридов , белков и липидов (так называемый липополисахарид). В клеточной стенке имеются поры диаметром 1-6 нм, которые делают её проницаемой для ряда молекул.
У грамотрицательных бактерий пептидогликановый слой неплотно прилегает к ЦПМ и имеет толщину лишь 2-3 нм. Он окружён наружной мембраной, имеющей, как правило, неровную, искривлённую форму. Между ЦПМ, слоем пептидогликана и внешней мембраной имеется пространство, называемое периплазматическим и заполненное раствором, включающим в себя транспортные белки и ферменты .
С внешней стороны от клеточной стенки может находиться капсула - аморфный слой, сохраняющий связь со стенкой. Слизистые слои не имеют связи с клеткой и легко отделяются, чехлы же не аморфны, а имеют тонкую структуру. Однако между этими тремя идеализированными случаями есть множество переходных форм.
Размеры
Размеры бактерий в среднем составляют 0,5-5 мкм . Масса - 4⋅10 −13 г . Escherichia coli , например, имеет размеры 0,3-1 на 1-6 мкм , Staphylococcus aureus - диаметр 0,5-1 мкм , Bacillus subtilis - 0,75 на 2-3 мкм . Крупнейшей из известных бактерий является Thiomargarita namibiensis , достигающая размера в 750 мкм (0,75 мм ). Второй является Epulopiscium fishelsoni , имеющая диаметр 80 мкм и длину до 700 мкм и обитающая в пищеварительном тракте хирурговой рыбы Acanthurus nigrofuscus . Achromatium oxaliferum достигает размеров 33 на 100 мкм , Beggiatoa alba - 10 на 50 мкм . Спирохеты могут вырастать в длину до 250 мкм при толщине 0,7 мкм . В то же время к бактериям относятся самые мелкие из имеющих клеточное строение организмов. Mycoplasma mycoides имеет размеры 0,1-0,25 мкм , что соответствует размеру крупных вирусов , например, табачной мозаики , коровьей оспы или гриппа . По теоретическим подсчётам, сферическая клетка диаметром менее 0,15-0,20 мкм становится неспособной к самостоятельному воспроизведению, поскольку в ней физически не могут поместиться все необходимые биополимеры и структуры в достаточном количестве.
При линейном увеличении радиуса клетки её поверхность возрастает пропорционально квадрату радиуса, а объём - пропорционально кубу, поэтому у мелких организмов отношение поверхности к объёму выше, чем у более крупных, что означает для первых более активный обмен веществ с окружающей средой. Метаболическая активность, измеренная по разным показателям, на единицу биомассы у мелких форм выше, чем у крупных. Поэтому небольшие даже для микроорганизмов размеры дают бактериям и археям преимущества в скорости роста и размножения по сравнению с более сложноорганизованными эукариотами и определяют их важную экологическую роль.
Многоклеточность у бактерий
Многоклеточный организм должен отвечать следующим условиям:
- его клетки должны быть агрегированы,
- между клетками должно осуществляться разделение функций,
- между агрегированными клетками должны устанавливаться устойчивые специфические контакты.
Многоклеточность у прокариот известна, наиболее высокоорганизованные многоклеточные организмы принадлежат к группам цианобактерий и актиномицетов . У нитчатых цианобактерий описаны структуры в клеточной стенке, обеспечивающие контакт двух соседних клеток - микроплазмодесмы . Показана возможность обмена между клетками веществом (красителем) и энергией (электрической составляющей трансмембранного потенциала). Некоторые из нитчатых цианобактерий содержат помимо обычных вегетативных клеток функционально дифференцированные: акинеты и гетероцисты . Последние осуществляют фиксацию азота и интенсивно обмениваются метаболитами с вегетативными клетками.
Способы передвижения и раздражимость
Многие бактерии подвижны. Имеется несколько принципиально различных типов движения бактерий. Наиболее распространено движение при помощи жгутиков: одиночных бактерий и бактериальных ассоциаций (роение). Частным случаем этого также является движение спирохет , которые извиваются благодаря аксиальным нитям, близким по строению к жгутикам , но расположенным в периплазме. Другим типом движения является скольжение бактерий , не имеющих жгутиков, по поверхности твёрдых сред и движение в воде безжгутиковых бактерий рода Synechococcus . Его механизм пока недостаточно изучен; предполагается участие в нём выделения слизи (проталкивание клетки) и находящихся в клеточной стенке фибриллярных нитей, вызывающих «бегущую волну» по поверхности клетки. Наконец, бактерии могут всплывать и погружаться в жидкости, меняя свою плотность, наполняя газами или опустошая аэросомы .
Бактерии активно передвигаются в направлении, определяемом теми или иными раздражителями. Это явление получило название таксис . Различают хемотаксис, аэротаксис, фототаксис и др.
Метаболизм
Конструктивный метаболизм
За исключением некоторых специфических моментов биохимические пути, по которым осуществляется синтез белков , жиров , углеводов и нуклеотидов , у бактерий схожи с таковыми у других организмов. Однако по числу возможных вариантов этих путей и, соответственно, по степени зависимости от поступления органических веществ извне они различаются.
Часть из них может синтезировать все необходимые им органические молекулы из неорганических соединений (автотрофы), другие же требуют готовых органических соединений, которые они способны лишь трансформировать (гетеротрофы).
Удовлетворять потребности в азоте бактерии могут как за счёт его органических соединений (подобно гетеротрофным эукариотам), так и за счёт молекулярного азота (как и некоторые археи). Большинство бактерий используют для синтеза аминокислот и других азотсодержащих органических веществ неорганические соединения азота: аммиак (поступающий в клетки в виде ионов аммония), нитриты и нитраты (которые предварительно восстанавливаются до ионов аммония). Фосфор они способны усваивать в виде фосфата , серу - в виде сульфата или реже сульфида .
Энергетический метаболизм
Способы же получения энергии у бактерий отличаются своеобразием. Существует три вида получения энергии (и все три известны у бактерий): брожение, дыхание и фотосинтез.
Бактерии, осуществляющие только бескислородный фотосинтез, не имеют фотосистемы II . Во-первых, это пурпурные и зелёные нитчатые бактерии, у которых функционирует только циклический путь переноса электронов, направленный на создание трансмембранного протонного градиента, за счёт которого синтезируется АТФ (фотофосфорилирование), а также восстанавливается НАД(Ф) + , использующийся для ассимиляции CO 2 . Во-вторых, это зелёные серные и гелиобактерии, имеющие и циклический, и нециклический транспорт электронов, что делает возможным прямое восстановление НАД(Ф) + . В качестве донора электрона, заполняющего «вакансию» в молекуле пигмента в бескислородном фотосинтезе используются восстановленные соединения серы (молекулярная, сероводород, сульфит) или молекулярный водород.
Существуют также бактерии с весьма специфическим энергетическим метаболизмом. Так, в октябре 2008 года в журнале Science появилось сообщение об обнаружении экосистемы, состоящей из представителей одного единственного ранее неизвестного вида бактерии - Desulforudis audaxviator , которые получают энергию для своей жизнедеятельности из химических реакций с участием водорода, образующегося в результате распада молекул воды под воздействием радиации залегающих вблизи нахождения колонии бактерий урановых руд . Некоторые колонии бактерий, обитающие на дне океана, используют для передачи энергии своим собратьям электрический ток .
Типы жизни
Объединить типы конструктивного и энергетического метаболизма можно в следующей таблице:
| Способы существования живых организмов (матрица Львова) | ||||
|---|---|---|---|---|
| Источник энергии | Донор электрона | Источник углерода | Название способа существования | Представители |
| ОВР | Неорганические соединения | Углекислый газ | Хемолитоавтотрофия | Нитрифицирующие, тионовые, ацидофильные железобактерии |
| Органические соединения | Хемолитогетеротрофия | Метанообразующие архебактерии, водородные бактерии | ||
| Органические вещества | Углекислый газ | Хемоорганоавтотрофия | Факультативные метилотрофы , окисляющие муравьиную кислоту бактерии | |
| Органические соединения | Хемоорганогетеротрофия | Большинство прокариот, из эукариот: животные , грибы , человек | ||
| Свет | Неорганические соединения | Углекислый газ | Фотолитоавтотрофия | Цианобактерии , пурпурные , зелёные бактерии , из эукариот: растения |
| Органические соединения | Фотолитогетеротрофия | Некоторые цианобактерии, пурпурные, зелёные бактерии | ||
| Органические вещества | Углекислый газ | Фотоорганоавтотрофия | Некоторые пурпурные бактерии | |
| Органические вещества | Фотоорганогетеротрофия | Галобактерии, некоторые цианобактерии , пурпурные, зелёные бактерии | ||
Из таблицы видно, что разнообразие типов питания прокариот гораздо больше, чем у эукариот (последние способны лишь к хемоорганогетеротрофии и фотолитоавтотрофии).
Размножение и устройство генетического аппарата
Размножение бактерий
Некоторые бактерии не имеют полового процесса и размножаются лишь равновеликим бинарным поперечным делением или почкованием . Для одной группы одноклеточных цианобактерий описано множественное деление (ряд быстрых последовательных бинарных делений, приводящий к образованию от 4 до 1024 новых клеток). Для обеспечения необходимой для эволюции и приспособления к изменчивой окружающей среде пластичности генотипа у них существуют иные механизмы.
Генетический аппарат
Гены, необходимые для жизнедеятельности и определяющие видовую специфичность, расположены у бактерий чаще всего в единственной ковалентно замкнутой молекуле ДНК - хромосоме (иногда для обозначения бактериальных хромосом, чтобы подчеркнуть их отличия от эукариотических, используют термин генофор (англ. genophore )). Область, где локализована хромосома, называется нуклеоид и не окружена мембраной. В связи с этим новосинтезированная мРНК сразу доступна для связывания с рибосомами, а транскрипция и трансляция сопряжены.
Отдельная клетка может содержать лишь 80 % от суммы генов, имеющихся во всех штаммах её вида (т. н. «коллективный геном»).
Помимо хромосомы, в клетках бактерий часто находятся плазмиды - также замкнутые в кольцо ДНК, способные к независимой репликации . Они могут быть настолько велики, что становятся неотличимы от хромосомы, но содержат дополнительные гены, необходимые лишь в специфических условиях. Специальные механизмы распределения обеспечивают сохранение плазмиды в дочерних клетках так, что они теряются с частотой менее 10 −7 в пересчёте на клеточный цикл. Специфичность плазмид может быть весьма разнообразной: от присутствия лишь у одного вида-хозяина до плазмиды RP4, встречающейся почти у всех грамотрицательных бактерий. В плазмидах кодируются механизмы устойчивости к антибиотикам , разрушения специфических веществ и т. д., nif-гены, необходимые для азотфиксации, также находятся в плазмидах. Ген плазмиды может включаться в хромосому с частотой около 10 −4 - 10 −7 .
В ДНК бактерий, как и в ДНК других организмов, выделяются транспозоны - мобильные сегменты, способные перемещаться из одной части хромосомы к другой, или во внехромосомные ДНК. В отличие от плазмид, они неспособны к автономной репликации и содержат IS-сегменты - участки, которые кодируют свой перенос внутри клетки. IS-сегмент может выступать в роли отдельной транспозоны.
Горизонтальный перенос генов
У прокариот может происходить частичное объединение геномов. При конъюгации клетка-донор в ходе непосредственного контакта передаёт клетке-реципиенту часть своего генома (в некоторых случаях весь). Участки ДНК донора могут обмениваться на гомологичные участки ДНК реципиента. Вероятность такого обмена значима только для бактерий одного вида.
Аналогично бактериальная клетка может поглощать и свободно находящуюся в среде ДНК, включая её в свой геном в случае высокой степени гомологии с собственной ДНК. Данный процесс носит название трансформация . В природных условиях протекает обмен генетической информацией при помощи умеренных фагов (трансдукция). Кроме этого, возможен перенос нехромосомных генов при помощи плазмид определённого типа, кодирующих этот процесс, процесс обмена другими плазмидами и передачи транспозон.
При горизонтальном переносе новых генов не образуется (как то имеет место при мутациях), однако осуществляется создание разных генных сочетаний. Это важно по той причине, что естественный отбор действует на всю совокупность признаков организма.
Клеточная дифференциация
Клеточная дифференциация - изменение набора белков (обычно также проявляющееся в изменении морфологии) при неизменном генотипе.
Образование покоящихся форм
Образование особо устойчивых форм с замедленным метаболизмом, служащих для сохранения в неблагоприятных условиях и распространения (реже для размножения) является наиболее распространённым видом дифференциации у бактерий. Наиболее устойчивыми из них являются эндоспоры , формируемые представителями Bacillus , Clostridium , Sporohalobacter , Anaerobacter (образует 7 эндоспор из одной клетки и может размножаться с их помощью ) и Heliobacterium . Образование этих структур начинается как обычное деление и на первых стадиях может быть превращено в него некоторыми антибиотиками. Эндоспоры многих бактерий способны выдерживать 10-минутное кипячение при 100 °C, высушивание в течение 1000 лет и, по некоторым данным, сохраняются в почвах и горных породах в жизнеспособном состоянии миллионы лет.
Менее устойчивыми являются экзоспоры , цисты (Azotobacter , скользящие бактерии и др.), акинеты (цианобактерии) и миксоспоры (миксобактерии).
Другие типы морфологически дифференцированных клеток
Актиномицеты и цианобактерии образуют дифференцированные клетки, служащие для размножения (споры, а также гормогонии и баеоциты соответственно). Необходимо также отметить структуры, подобные бактероидам клубеньковых бактерий и гетероцистам цианобактерий, служащие для защиты нитрогеназы от воздействия молекулярного кислорода.
Классификация
Наибольшую известность получила фенотипическая классификация бактерий, основанная на строении их клеточной стенки, включённая, в частности, в IX издание Определителя бактерий Берджи (1984-1987). Крупнейшими таксономическими группами в ней стали 4 отдела: Gracilicutes (грамотрицательные), Firmicutes (грамположительные), Tenericutes (микоплазмы) и Mendosicutes (археи).
В последнее время всё большее развитие получает филогенетическая классификация бактерий (и именно она используется в Википедии), основанная на данных молекулярной биологии. Одним из первых методов оценки родства по сходству генома был предложенный ещё в 1960-х годах метод сравнения содержания гуанина и цитозина в ДНК. Хотя одинаковые значения их содержания и не могут дать никакой информации об эволюционной близости организмов, их различия на 10 % означают, что бактерии не принадлежат к одному роду. Другим методом, произведшим в 1970-е настоящую революцию в микробиологии, стал анализ последовательности генов в 16s рРНК , который позволил выделить несколько филогенетических ветвей эубактерий и оценить связи между ними. Для классификации на уровне вида применяется метод ДНК-ДНК гибридизации . Анализ выборки хорошо изученных видов позволяет считать, что 70 % уровень гибридизации характеризует один вид, 10-60 % - один род, менее 10 % - разные роды.
Филогенетическая классификация отчасти повторяет фенотипическую, так, группа Gracilicutes присутствует и в той и в другой. В то же время систематика грамотрицательных бактерий была полностью пересмотрена, архебактерии и вовсе выделены в самостоятельный таксон высшего ранга , часть таксономических групп разбита на части и перегруппирована, в одни группы объединены организмы с совершенно разными экологическими функциями, что вызывает ряд неудобств и недовольство части научного сообщества. Объектом нареканий становится и то, что проводится фактически классификация молекул, а не организмов.
Происхождение, эволюция, место в развитии жизни на Земле
Бактерии наряду с археями были одними из первых живых организмов на Земле, появившись около 3,9-3,5 млрд лет назад. Эволюционные взаимоотношения между этими группами ещё до конца не изучены, есть как минимум три основные гипотезы : Н. Пэйс предполагает наличие у них общего предка протобактерии, Заварзин считает архей тупиковой ветвью эволюции эубактерий, освоившей экстремальные местообитания; наконец, по третьей гипотезе археи - первые живые организмы, от которых произошли бактерии.
Патогенные бактерии
Патогенными называются бактерии, паразитирующие на других организмах. Бактерии вызывают большое количество заболеваний человека, таких как чума (Yersinia pestis ), сибирская язва (Bacillus anthracis ), лепра (проказа, возбудитель: Mycobacterium leprae ), дифтерия (Corynebacterium diphtheriae ), сифилис (Treponema pallidum ), холера (Vibrio cholerae ), туберкулёз (Mycobacterium tuberculosis ), листериоз (Listeria monocytogenes ) и др. Открытие патогенных свойств у бактерий продолжается: в 1976 обнаружена болезнь легионеров , вызываемая Legionella pneumophila , в 1980-е -1990-е годы было показано, что Helicobacter pylori вызывает язвенную болезнь и даже рак желудка , а также хронический
Бактерии это одноклеточные организмы, лишенные хлорофилла.
Бактерии встречаются повсеместно, населяя все среды обитания. Наибольшее количество их находится в почве на глубине до 3 км (до 3 миллиардов в одном грамме почвы). Их много в воздухе (на высоте до 12 км), в организмах животных и растений (как живых, так и мертвых), не является исключением и организм человека.
Среди бактерий встречаются неподвижные и подвижные формы. Передви-гаются бактерии с помощью одного или нескольких жгути-ков, которые располагаются на всей поверхности тела или на определенном участке.
Клетки бактерий разнообразны по форме:
- шаровидные - кокки,
- палочковидные - ба-циллы,
- в форме запятой - вибрионы,
- извитые - спириллы.
Кокки :
Монококки: это отдельно расположенные клетки.
Диплококки: это парные кокки, после деления могут образовывать пары.
Гонококк Нейссера: возбудитель гонореи
Пневмококки: возбудитель крупозной пневмонии
Менингококки: возбудитель менингита (острое воспаление мозговых оболочек)
Стрептококки: это клетки округлой формы, которые после деления образуют цепочки.
α - зеленящие стрептококки
β - гемолитические стрептококки возбудители скарлатины, ангины, фарингита…
γ - не гемолитические стрептококки
Стафилококки: это группа микроорганизмов, которая не разошлась после деления, образует огромные беспорядочные грозди.
Возбудитель: гнойничковых заболеваний, сепсиса, фурункулов, абсцессов, флегмон, мастита, пиодермита и пневмонию у новорожденных.
Сарцины: это скопление кокков в группы в виде пакетиков по 8 и более кокков.
Палочковидные:
Это бактерии цилиндрической формы, похожие на палочки размером 1-5×0,5-1 мкм, чаще располагаются одиночно.
Собственно бактерии: это палочковидные бактерии, которые не образуют споры.
Бациллы: это палочковидные бактерии, которые образуют споры.
(бацилла Коха, кишечная палочка, возбудитель сибирской язвы, синегнойная палочка, возбудитель чумы, возбудитель коклюша, возбудитель мягкого шанкра, возбудитель столбняка, возбудитель ботулизма, возбудитель…)
Вибрионы:
Это слабо изогнутые клетки, напоминающие по форме запятые размером 1-3 мкм.
Холерный вибрион: возбудитель холеры. Обитает в воде, через которую происходит заражение.
Спириллы:
Это извитые микроорганизмы в виде спирали, с одни, двумя и более спиралевидными кольцами.
Безвредные бактерии, живущие в сточных водах и запруженных водоемах.
Спирохеты:
Это тонкие длинные топоровидные бактерии, представлены тремя видами: Трепонемы, Боррелия, Лертоспира. Для человека патогенна бледная трепонема - возбудитель сифилива передается половым путем.
Строение бактериальной клетки:
Структура бактериальной клетки хорошо изучена с помощью электронной микроскопии. Бактериальная клетка состоит из оболочки, наружный слой которой называется клеточная стенка, а внутренний - цитоплазматическая мембрана, а также цитоплазмы с включениями и нуклеотидами. Имеются дополнительные структуры: капсула, микрокапсула, слизь, жгутики, пили, плазмиды;
Клеточная стенка - прочная, упругая структура, придающая бактерии определенную форму, и «сдерживающая» высокое осмотическое давление в бактериальной клетке. Она защищает клетку от дейст-вия вредных факторов внешней среды.
Наружная мембрана представлена липополисахаридами, фосфолипидами и белками. С ее внешней стороны расположен липо-полисахарид.
Между клеточной стенкой и цитоплазматической мембранами находится периплазматическое пространство, или периплазма, содержащая ферменты.
Цитоплазматическая мембрана прилегает к внутренней по-верхности клеточной стенки бактерий и окружает наружную часть цитоплазмы бактерий. Она состоит из двойного слоя липидов, а также интегральных белков, пронизывающих ее насквозь.
Цитоплазма занимает основной объем бактериальной клетки и состоит из растворимых белков, рибонуклеиновых кислот, включений и многочисленных мелких гранул - рибосом, ответст-венных за синтез белков. В цитоплазме имеются различные включения в виде гранул гликогена, полисахаридов, жирных ки-слот и полифосфатов.
Нуклеотид - эквивалент ядра у бактерий. Он расположен в цито-плазме бактерий в виде двух нитчатой ДНК, замкнутой в кольцо и плотно уложенной наподобие клубка. Обычно в бактериальной клетке содер-жится одна хромосома, представленная замкнутой в кольцо мо-лекулой ДНК.
Кроме нуклеотида в бактериальной клетке могут находиться внехромосомные факторы наследственности - плазмиды, пред-ставляющие собой ковалентно замкнутые кольца ДНК и способ-ные к репликации независимо от бактериальной хромосомы.
Капсула - слизистая структура, прочно связанная с клеточной стенкой бактерий и имеющая четко очерченные внешние грани-цы. Обычно капсула состоит из полисахаридов, иногда из поли-пептидов,
Многие бактерии содержат микрокапсулу - слизистое образова-ние, выявляемое лишь при электронной микроскопии.
Жгутики бактерий определяют подвижность клетки. Жгутики представляют собой тонкие нити, берущие начало от цитоплазматической мембраны, они прикреплены к цитоплазматической мембране и клеточной стенке специальными дисками, имеют большую длину, они состоят из белка - флагеллина, закрученного в виде спирали. Жгутики выяв-ляют с помощью электронного микроскопа.
Споры - своеобразная форма покоящихся грамположительных бактерий, образующихся во внешней среде при неблагопри-ятных условиях существования бактерий (высушивание, дефицит питательных веществ и др.).
L-формы бактерий .
У многих бактерий при частичном или полном разрушении клеточных стенок образуются L-формы. У некоторых они возникают спонтанно. Обра-зование L-форм происходит под действием пенициллина, который нарушает синтез мукопептидов клеточной стен-ки. По морфологии L-формы разных видов бактерий сходны между собой. Они представляют шаровидные, образования различной величины: от 1-8 мкм до 250 нм, они способных, как и вирусы, прохо-дить через поры фарфоровых фильтров. Однако в отли-чие от вирусов L-формы можно выращивать на искусст-венных питательных средах, добавляя к ним пенициллин, сахара, лошадиную сыворотку. При удалении из пита-тельной среды пенициллина L-формы вновь превращают-ся в исходные формы бактерий.
В настоящее время получены L-формы протея, кишечной палочки, холерного вибриона, бруцелл, возбудителей газовой гангрены и столбняка и других микроорганизмов.
Грамположительные микроорганизмы (гр + м/о).
К ним относят : золотистый и эпидермальный стафилококк и стрептококк...
Место обитания : верхние дыхательные пути и кожа.
Резервуар : кожа, воздух, предметы ухода, мебель, постельные принадлежности, одежду.
При высушивании не погибают.
Размножение: вне человека не размножаются, но способны к размножению в продуктах питания при не правильном хранении.
Грамотрицательные микроорганизмы (гр - м /о).
К ним относят : кишечная палочка, клебсиелла, цитробактер, протей, синегнойная палочка...
Место обитания : кишечник, слизистая мочевыводящих и дыхательных путей…
Резервуар : влажная ветошь, щетки для мытья посуды, дыхательная аппаратура, влажные поверхности, лекарственные и слабые дез. растворы.
При высушивании погибают.
Размножение: накапливаются во внешней среде, в дез. растворах с заниженной концентрацией.
Передаются : воздушно-капельным и контактно-бытовым путем.